
General Senses
Anatomy & Physiology I
Is the water is the shower the right temperature? Ouch what stung me? We ask ourselves questions like this daily, but how do we know the answers? Being able to sense external stimuli, interpret, and respond accordingly is critical for continued survival; and is made possible by your nervous system and the process of sensation. Sensation is defined as a perception or feeling that results from a stimulus. This stimulus will come in a variety of forms and could arise from either the external or internal environment. We know this verbose definition of sensation as simply our senses.
If you were to ask someone what the senses are, and they are likely to list the five major senses—taste, smell, touch, hearing, and sight. However, these are not all of the senses. The most obvious omission from this list is equilibrium. Also, what is referred to simply as touch can be further subdivided into pressure, vibration, itch, tickle, and hair-follicle position, on the basis of the type of receptors that perceive these touch sensations. Other overlooked senses include temperature perception, pain perception, and proprioception.
Senses can be classified as either general or specific. A general sense utilizes relatively simple receptors that are widely distributed throughout the body; meaning that you can tell the temperature of the water in the shower with your hand, foot, arm, face, etc. General senses often contribute to the sense of touch (as described above), or to proprioception (body position in space) and kinesthesia (body movement), or to a visceral sense (internal environment monitoring that stimulates the ANS). A special sense utilizes complex receptors and has a specific organ devoted to it, namely the eye, inner ear, tongue, or nose. This means that you can only see with your eye, not the back of your head (sorry mom) because there are no receptors for light there. This module focuses on general senses, the special sense will be addressed in a later module. A quick comparison between general and special senses is listed below.
|
General Senses |
Special Senses |
|
Simple receptor and afferent pathway |
Complex receptor and afferent pathway |
|
Widely distributed all over the body |
Concentrated in the head and face |
|
Includes: pressure, pain, itch, tickle, vibration, temperature, and proprioception. |
Includes: vision, olfaction (smell), gustation (taste), audition (hearing), and equilibrium. |
Each of the senses is referred to as a sensory modality - modality refers to the way that information is encoded. Each modality has a specific receptor that is activated by a particular stimulus and a specific sensory neuron that carries the information from the receptor into the CNS. If you remember back to the module that addressed homoeostasis there was a flow of information from receptor into the CNS and then back out to the effector. The receptor senses the change in the external environment. This sensory information is then passed up the afferent pathway via a unipolar neuron into the posterior grey horn of the spinal cord. An interneuron then processes the sensory information deciding how to respond and creating a motor signal. This motor output is then passes down the efferent pathway via a multipolar neuron form the anterior grey horn of the spinal cord to the effector. The effector can then act allowing us to respond to the stimulus that set this chain of events in motion. See the diagram below for a visual representation of this pathway.
Figure 1: Testing the water: (1) the sensory neuron has endings in the skin that sense a stimulus such as water temperature. The strength of the signal that starts here is dependent on the strength of the stimulus. (2) The graded potential from the sensory endings, if strong enough, will initiate an action potential at the initial segment of the axon (which is immediately adjacent to the sensory endings in the skin). (3) The axon of the peripheral sensory neuron enters the spinal cord and contacts another neuron in the gray matter. The contact is a synapse where another graded potential is caused by the release of a chemical signal from the axon terminals. (4) An action potential is initiated at the initial segment of this neuron and travels up the sensory pathway to a region of the brain called the thalamus. Another synapse passes the information along to the next neuron. (5) The sensory pathway ends when the signal reaches the cerebral cortex. (6) After integration with neurons in other parts of the cerebral cortex, a motor command is sent from the precentral gyrus of the frontal cortex. (7) The upper motor neuron sends an action potential down to the spinal cord. The target of the upper motor neuron is the dendrites of the lower motor neuron in the gray matter of the spinal cord. (8) The axon of the lower motor neuron emerges from the spinal cord in a nerve and connects to a muscle through a neuromuscular junction to cause contraction of the target muscle. CCBY: OpenStax college
View "General Senses" on YouTube (opens in a new window)
Helpful Study Fact #1: Words that go together – when you see one of these words being used to reference the general senses you know that the other 3 will soon follow. Group 1: sensory information, afferent pathway, unipolar neuron, and posterior/dorsal. Group 2: motor information, efferent pathway, multipolar neuron, and anterior/ventral.
The somatic nervous system is responsible for our conscious perception of the environment and for our voluntary responses to that perception by means of skeletal muscle movement. Peripheral sensory neurons receive input from environmental stimuli, but the neurons that produce motor responses originate in the central nervous system. Let's now look at a simple reflex action - the withdrawal reflex. When you touch a hot stove, you pull your hand away. Sensory receptors in the skin sense extreme temperature and the early signs of tissue damage (see section on selectivity – insert link to that portion of the module). This triggers an action potential, which travels along the sensory fiber from the skin, through the dorsal spinal root to the spinal cord, and directly activates a ventral horn motor neuron. That neuron sends a signal along its axon to excite the biceps brachii, causing contraction of the muscle and flexion of the forearm at the elbow to withdraw the hand from the hot stove. This basic withdrawal reflex includes sensory input (the painful stimulus), central processing (the synapse in the spinal cord), and motor output (activation of a ventral motor neuron that causes contraction of the biceps brachii).

Describe the locations and functions of receptors for tactile, thermal, and pain sensations, and for proprioception.
Now that we have seen the pathway in its entirety let's look closely at the beginning of the process of sensation, the sensory receptors. Receptors are the cells or structures that detect sensations because a receptor is changed directly by a stimulus. Stimuli in the environment activate specialized receptor cells in the peripheral nervous system. Different types of stimuli are sensed by different types of receptor cells. Meaning that the major role of sensory receptors is to help us learn about the environment around us, or about the state of our internal environment. Receptors fall into two main categories: receptors that are standalone cells or those that are part of the unipolar sensory neuron. A great example of a stand-alone receptor cells is a touch receptor in the skin known as a Merkel's disc. As you can see from the figure below the receptor is not directly connected to the sensory neuron but rather they are closely associated so that when the signal can be passed from the receptor to the sensory neuron efficiently. The other category of receptor can be seen in the case of pain receptors (nociceptors). As seen below, the nociceptors are part of the sensory neuron and when stimulated the signal travels up the neuron into the CNS.

Figure 2: To the left of the sectioned hair shaft you can see a yellow largely diffuse neuron, this is a free nerve ending. Notice how the neuron simply terminates in the tissue and serves as a receptor in that location. The box to the right identifies a Merkel's disc. Notice how this receptor cell is separate from the yellow neuron and stands alone. CCBY: Blausen.com staff.
Now to name the many different types of receptors we use three different criteria: cell type, position, and function. Receptors can be classified structurally on the basis of cell type and their position in relation to stimuli they sense. They can also be classified functionally on the basis of the transduction of stimuli, or how the mechanical stimulus, light, or chemical changed the cell membrane potential.
As mentioned before, the cells that interpret information about the environment can be either (1) a specialized receptor cell, which has distinct structural components that interpret a specific type of stimulus; or (2) neuron that has an encapsulated ending or free nerve ending with dendrites embedded in tissue that would receive a sensation (see the figure below). The pain and temperature receptors in the dermis of the skin are examples of neurons that have free nerve endings. Also located in the dermis of the skin are lamellated corpuscles, neurons with encapsulated nerve endings that respond to pressure and touch. The cells in the retina that respond to light stimuli are an example of a specialized receptor, a photoreceptor.

Figure 3: Receptor cell types can be classified on the basis of their structure. Sensory neurons can have either (a) free nerve endings or (b) encapsulated endings. Photoreceptors in the eyes, such as rod cells, are examples of (c) specialized receptor cells. These cells release neurotransmitters onto a bipolar cell, which then synapses with the optic nerve neurons.
Another way that receptors can be classified is based on their location relative to the stimuli. An exteroceptor is a receptor that is located near a stimulus in the external environment, such as the somatosensory receptors that are located in the skin. An interoceptor is one that interprets stimuli from internal organs and tissues, such as the receptors that sense the increase in blood pressure in the aorta or carotid sinus. Finally, a proprioceptor is a receptor located near a moving part of the body, such as a muscle, that interprets the positions of the tissues as they move.
Functionally, receptor cells can be further categorized on the basis of the type of stimuli they transduce. Chemical stimuli can be interpreted by a chemoreceptor that interprets chemical stimuli, such as an object's taste or smell. Osmoreceptors respond to solute concentrations of body fluids. Additionally, pain is primarily a chemical sense that interprets the presence of chemicals from tissue damage, or similar intense stimuli, through a nociceptor. Physical stimuli, such as pressure and vibration, as well as the sensation of sound and body position (balance), are interpreted through a mechanoreceptor. Another physical stimulus that has its own type of receptor is temperature, which is sensed through a thermoreceptor that is either sensitive to temperatures above (heat) or below (cold) normal body temperature.
Stimuli from varying sources, and of different types, are received and changed into the electrochemical signals of the nervous system. This occurs when a stimulus changes the cell membrane potential of a sensory neuron. The stimulus causes the sensory cell to produce a graded potential that is called a receptor potential if the receptor is a stand-alone cell or a generator potential if the receptor is a part of the sensory neuron. This graded potential then creates an action potential that is relayed into the central nervous system (CNS), where it is integrated with other sensory information—or sometimes higher cognitive functions—to become a conscious perception of that stimulus. The central integration may then lead to a motor response.
Somatosensation is the group of sensory modalities that are associated with touch and proprioception. These modalities include pressure, vibration, light touch, tickle, itch, temperature, pain, and proprioception. This means that its receptors are not associated with a specialized organ, but are instead spread throughout the body in a variety of organs. Many of the somatosensory receptors are located in the skin, but receptors are also found in muscles, tendons, joint capsules, ligaments, and in the walls of visceral organs.
Two types of somatosensory signals that are transduced by free nerve endings are pain and temperature. These two modalities use thermoreceptors and nociceptors to transduce temperature and pain stimuli, respectively. Temperature receptors are stimulated when local temperatures differ from body temperature. Some thermoreceptors are sensitive to just cold and others to just heat. Nociception is the sensation of potentially damaging stimuli. Mechanical, chemical, or thermal stimuli beyond a set threshold will elicit painful sensations. Stressed or damaged tissues release chemicals that activate receptor proteins in the nociceptors. For example, the sensation of heat associated with spicy foods involves capsaicin, the active molecule in hot peppers. Capsaicin molecules bind to a specific ion channel on the nociceptor, one that is sensitive to temperatures above 37°C. The dynamics of capsaicin binding with a particular ion channel is unusual in that the molecule remains bound for a long time. Because of this, it will decrease the ability of other stimuli to elicit pain sensations through the activated nociceptor. For this reason, capsaicin can be used as a topical analgesic, such as in products such as Icy Hot™.
If you drag your finger across a textured surface, the skin of your finger will vibrate. Such low frequency vibrations are sensed by mechanoreceptors called Merkel cells, also known as type I cutaneous mechanoreceptors. Merkel cells are located in the stratum basale of the epidermis. Deep pressure and vibration is transduced by lamellated (Pacinian) corpuscles, which are receptors with encapsulated endings found deep in the dermis, or subcutaneous tissue. Light touch is transduced by the encapsulated endings known as tactile (Meissner) corpuscles. Follicles are also wrapped in a plexus of nerve endings known as the hair follicle plexus. These nerve endings detect the movement of hair at the surface of the skin, such as when an insect may be walking along the skin. Stretching of the skin is transduced by stretch receptors known as bulbous corpuscles. Bulbous corpuscles are also known as Ruffini corpuscles, or type II cutaneous mechanoreceptors.
Other somatosensory receptors are found in the joints and muscles. Stretch receptors monitor the stretching of tendons, muscles, and the components of joints. For example, have you ever stretched your muscles before or after exercise and noticed that you can only stretch so far before your muscles spasm back to a less stretched state? This spasm is a reflex that is initiated by stretch receptors to avoid muscle tearing. Such stretch receptors can also prevent over-contraction of a muscle. In skeletal muscle tissue, these stretch receptors are called muscle spindles. Tendon organs similarly transduce the stretch levels of tendons The types of nerve endings, their locations, and the stimuli they transduce are presented in the table.
|
* No corresponding eponymous name. |
|||
|
Mechanoreceptors of Somatosensation |
|||
|
Name |
Historical (eponymous) name |
Location(s) |
Stimuli |
|
Free nerve endings |
* |
Dermis, cornea, tongue, joint capsules, visceral organs |
Pain, temperature, mechanical deformation |
|
Mechanoreceptors |
Merkel's discs |
Epidermal–dermal junction, mucosal membranes |
Low frequency vibration (5–15 Hz) |
|
Bulbous corpuscle |
Ruffini's corpuscle |
Dermis, joint capsules |
Stretch |
|
Tactile corpuscle |
Meissner's corpuscle |
Papillary dermis, especially in the fingertips and lips |
Light touch, vibrations below 50 Hz |
|
Lamellated corpuscle |
Pacinian corpuscle |
Deep dermis, subcutaneous tissue |
Deep pressure, high-frequency vibration (around 250 Hz) |
|
Hair follicle plexus |
* |
Wrapped around hair follicles in the dermis |
Movement of hair |
|
Muscle spindle |
* |
In line with skeletal muscle fibers |
Muscle contraction and stretch |
|
Tendon stretch organ |
Golgi tendon organ |
In line with tendons |
Stretch of tendons |
CCBY: OpenStax college

Once any sensory cell transduces (converts) a stimulus into a nerve impulse, that impulse has to travel along sensory axons to reach the CNS. The nerves that convey sensory information from the periphery to the CNS are either spinal nerves, connected to the spinal cord, or cranial nerves, connected to the brain.
Generally, spinal nerves contain afferent axons from sensory receptors in the periphery, such as from the skin, mixed with efferent axons travelling to the muscles or other effector organs. As the spinal nerve nears the spinal cord, it splits into dorsal and ventral roots. The dorsal root contains only the axons of sensory neurons, whereas the ventral roots contain only the axons of the motor neurons. (Remember the words that go together section we already covered?) Typically, spinal nerve systems that connect to the brain are contralateral, in that the right side of the body is connected to the left side of the brain and the left side of the body to the right side of the brain. This crossing over of information from right body to left brain is termed decussation and can happen at two locations: the pyramids of the medulla oblongata and in the white matter of the spinal cord. In the simplest terms, the right hemisphere of the brain receives information from and controls motor output to the left side of the body, and vice versa for the left hemisphere of the brain. See the diagram below for a visual depiction of decussation.
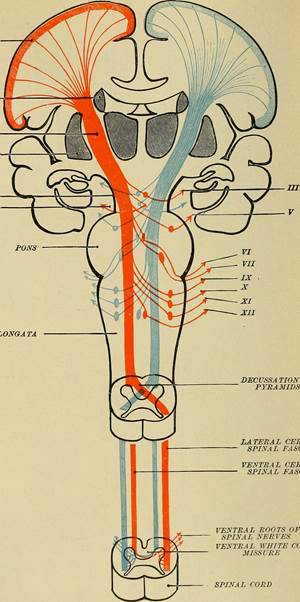
Figure 4: Notice how the blue information originates and enters on the left side of the spinal cord and as it ascends, will decussate (cross over/switch side) at the pyramids of the medulla. This decussation means that information originating on the left side of the body is interpreted by the right side of the brain. The opposite is true is and illustrated by the orange lines. CCBY: Flickr API
Cranial nerves convey specific sensory information from the head and neck directly to the brain. Whereas spinal information is contralateral, cranial nerve systems are mostly ipsilateral, meaning that a cranial nerve on the right side of the head is connected to the right side of the brain. Some cranial nerves contain only sensory axons, such as the olfactory, optic, and vestibulocochlear nerves. Other cranial nerves contain both sensory and motor axons, including the trigeminal, facial, glossopharyngeal, and vagus nerves (however, the vagus nerve is not associated with the somatic nervous system). The general senses of somatosensation for the face travel through the trigeminal system.
.
A sensory pathway that carries peripheral sensations to the brain is referred to as an ascending pathway, or ascending tract. The various sensory modalities each follow specific pathways through the CNS. Somatosensory stimuli from below the neck pass along the sensory pathways of the spinal cord, whereas somatosensory stimuli from the head and neck travel through the cranial nerves. The somatosensory pathways are divided into two separate systems on the basis of the location of the receptor neurons. The dorsal column system (also referred to as the medial lemniscus pathway) and the spinothalamic tract are the two major pathways that bring sensory information to the brain. The sensory pathways in each of these systems are composed of three successive neurons.
The dorsal column system begins with the axon of a dorsal root ganglion neuron entering the dorsal root and joining the dorsal column white matter in the spinal cord. This means that the dendrites of the first neuron in the dorsal column system receive the signal from the stimulated receptor. The action potential travels up the dendrites, into the spinal nerve, into the dorsal root, and then finally reaching the soma of the neuron in the dorsal root ganglion. Once the signal is processed by the soma of this unipolar neuron, the action potential travels down the axon from the dorsal root ganglion and into the posterior grey horn. The axon does not end here, but rather projects into the posterior white columns and ascends up the spinal cords into the medulla oblongata. Because this dorsal root ganglion neuron is the first to carry the information from receptor to higher processing in the cerebrum it is termed the first order neuron.
The next step in this system is for a multipolar neuron whose soma resides in the medulla, receives the information from the first order neuron and decussates – crosses the midline of the medulla - before continuing to ascend toward the cerebrum. Since this is the second neuron of the system, it is termed the second order neuron. Think of a relay race where one runner takes a lap around the track but then has to pass the baton to the next runner who will take their lap around the track. This is made possible because the first runner ends where the second runner begins, so the signal (the baton) can be passed efficiently from one runner to the next. This is very similar to how the somatosensory pathways operate. The first runner (first order neuron) brought the baton (signal) from the starting line (the receptor) into the medulla. This is where the second runner (second order neuron) picks up the baton and carries it toward the cerebrum to the next runner. These axons then continue to ascend the brain stem as a bundle called the medial lemniscus.
The second order neurons terminate in the thalamus, where each synapses with the third order neuron (third runner) in their respective pathway. The third neuron in the system projects its axons to the postcentral gyrus of the cerebral cortex also known as the primary sensory cortex, where somatosensory stimuli are initially processed and the conscious perception of the stimulus occurs.
The spinothalamic tract begins with dorsal root ganglion neurons, the same type of first order neuron that was used and described in the dorsal column system. However, the axons of these first order neurons do not ascend the spinal cord, but rather they terminate in the posterior grey horn. This is where the baton is passed from the first order neuron to the awaiting second order neuron. The name "spinothalamic" comes from this second neuron, which has its cell body in the spinal cord gray matter and connects to the thalamus. Axons from these second order neurons then decussate within the spinal cord and ascend to the brain and enter the thalamus, where each synapses with the third neuron in its own pathway.
These two systems are similar in that they both begin with dorsal root ganglion cells, as with most general sensory information. The dorsal column system is primarily responsible for touch sensations and proprioception, whereas the spinothalamic tract pathway is primarily responsible for pain and temperature sensations. Another similarity is that the second neurons in both of these pathways are contralateral, because they project across the midline to the other side of the brain or spinal cord. In the dorsal column system, this decussation takes place in the brain stem; in the spinothalamic pathway, it takes place in the spinal cord at the same spinal cord level at which the information entered. The third neurons in the two pathways are essentially the same, synapsing in the thalamus, and the thalamic neuron projects to the somatosensory cortex. See the figure below for a side by side illustration of both systems.
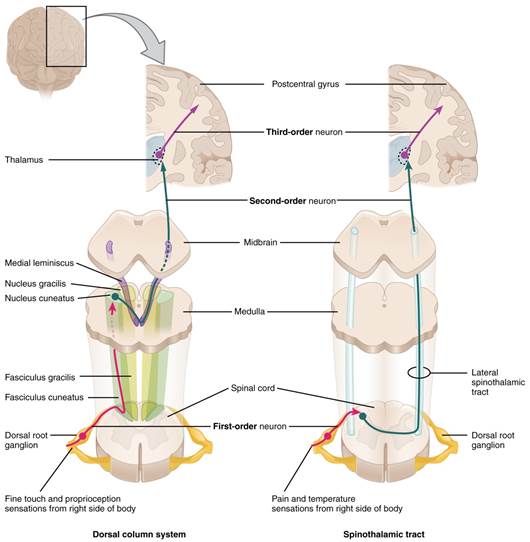
Figure 5: The dorsal column system and spinothalamic tract are the major ascending pathways that connect the periphery with the brain. CCBY: OpenStax college
The diencephalon is beneath the cerebrum and includes the thalamus and hypothalamus. In the somatic nervous system, the thalamus is an important relay for communication between the cerebrum and the rest of the nervous system. The hypothalamus has both somatic and autonomic functions. In addition, the hypothalamus communicates with the limbic system, which controls emotions and memory functions.
Sensory input to the thalamus comes from most of the special senses and ascending somatosensory tracts. The thalamus is a required transfer point for most sensory tracts that reach the cerebral cortex, where conscious sensory perception begins. The one exception to this rule is the olfactory system. The olfactory tract axons from the olfactory bulb project directly to the cerebral cortex. The thalamus is the phone operator of somatosensation. If you call a college campus but don't know the number for the bookstore, you will talk to the operator who will connect you to the right destination. The thalamus does this for sensory information; it takes the second order neuron, and says "How may I direct your call?" connecting it to the correct third order neuron so the information reaches the correct destination. The anterior thalamus serves as a relay between the hypothalamus and the emotion and memory-producing limbic system. The medial portion serves as a relay for information from the limbic system and basal ganglia to the cerebral cortex. This allows memory creation during learning, but also determines alertness. The special and somatic senses connect to the lateral portion, where their information is relayed to the appropriate sensory cortex of the cerebrum.
The cerebral cortex maintains the sensory topography of the body in reference to the position of the receptor cells. The somatosensory cortex (postcentral gyrus) provides an example in which, in essence, the locations of the somatosensory receptors in the body are mapped onto the somatosensory cortex. This mapping is often depicted using a sensory homunculus (see the figure below).
The term homunculus comes from the Latin word for "little man" and refers to a map of the human body that is laid across a portion of the cerebral cortex. In the somatosensory cortex, the external genitals, feet, and lower legs are represented on the medial face of the gyrus within the longitudinal fissure. As the gyrus curves out of the fissure and along the surface of the parietal lobe, the body map continues through the thighs, hips, trunk, shoulders, arms, and hands. The head and face are lateral to the fingers as the gyrus approaches the lateral sulcus. The representation of the body in this topographical map is medial to lateral from the lower to upper body. The connections through the thalamus maintain topography such that the anatomic information is preserved. Note that this correspondence does not result in a perfectly miniature scale version of the body, but rather exaggerates the more sensitive areas of the body, such as the fingers and lower face. Less sensitive areas of the body, such as the shoulders and back, are mapped to smaller areas on the cortex.
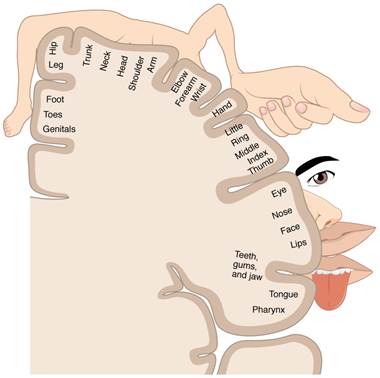
Figure 6: A cartoon representation of the sensory homunculus arranged adjacent to the cortical region in which the processing takes place. Note the exaggeration of the more sensitive body area ie. the size of the hand versus the elbow. CCBY: OpenStax college
Sensory input to the brain enters through pathways that travel through either the spinal cord (for somatosensory input from the body) or the brain stem (for everything else, except the visual and olfactory systems) to reach the diencephalon. In the diencephalon, sensory pathways reach the thalamus. This is necessary for all sensory systems to reach the cerebral cortex, except for the olfactory system that is directly connected to the frontal and temporal lobes.
The cortex has been described as having specific regions that are responsible for processing specific information; there is the visual cortex, somatosensory cortex, gustatory cortex, etc. However, our experience of these senses is not divided. Instead, we experience what can be referred to as a seamless percept. Our perceptions of the various sensory modalities—though distinct in their content—are integrated by the brain so that we experience the world as a continuous whole. In the cerebral cortex, sensory processing begins at the primary sensory cortex, then proceeds to an association area, and finally, into a multimodal integration area.
Continuing on, the sensory stimuli has been received by receptors and relayed to the CNS along ascending pathways. In the cerebral cortex, the initial processing of sensory perception progresses to incorporation of sensory perceptions into memory, and more importantly, they lead to a response. The completion of cortical processing in sensory areas initiates a similar progression of motor processing, usually in different cortical areas.
Whereas the sensory cortical areas are located in the occipital, temporal, and parietal lobes, motor functions are largely controlled by the frontal lobe. The most anterior regions of the frontal lobe—the prefrontal areas—are important for executive functions, which are those cognitive functions that lead to goal-directed behaviors. These higher cognitive processes include working memory, which has been called a "mental scratch pad," that can help organize and represent information that is not in the immediate environment. The prefrontal lobe is responsible for aspects of attention, such as inhibiting distracting thoughts and actions so that a person can focus on a goal and direct behavior toward achieving that goal.
The functions of the prefrontal cortex are integral to the personality of an individual, because it is largely responsible for what a person intends to do and how they accomplish those plans. A famous case of damage to the prefrontal cortex is that of Phineas Gage, dating back to 1848. He was a railroad worker who had a metal spike impale his prefrontal cortex (see the figure). He survived the accident, but according to second-hand accounts, his personality changed drastically. Friends described him as no longer acting like himself. Whereas he was a hardworking, amiable man before the accident, he turned into an irritable, temperamental, and lazy man after the accident. The accounts suggest that some aspects of his personality did change. (Later accounts from when he worked as a stage coach driver showed that some of the more sociable aspects of his personality did return.) (credit b: John M. Harlow, MD)
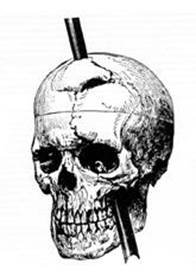
Figure 7: Phineas Gage, 1848, was a railroad worker who had a metal spike impale his prefrontal cortex.
In generating motor responses, the executive functions of the prefrontal cortex will need to initiate actual movements. One way to define the prefrontal area is any region of the frontal lobe that does not elicit movement when electrically stimulated. These are primarily in the anterior part of the frontal lobe. The regions of the frontal lobe that remain are the regions of the cortex that produce movement. The prefrontal areas project into the secondary motor cortices, which include the premotor cortex and the supplemental motor area. Two important regions that assist in planning and coordinating movements are located adjacent to the primary motor cortex. The premotor area aids in controlling movements of the core muscles to maintain posture during movement, whereas the supplemental motor area is hypothesized to be responsible for planning and coordinating movement. The supplemental motor area also manages sequential movements that are based on prior experience (that is, learned movements).
The primary motor cortex is located in the precentral gyrus of the frontal lobe. The primary motor cortex receives input from several areas that aid in planning movement, and its principle output stimulates spinal cord neurons to stimulate skeletal muscle contraction. The primary motor cortex is arranged in a similar fashion to the primary somatosensory cortex, in that it has a topographical map of the body, creating a motor homunculus (see the figure below). The neurons responsible for musculature in the feet and lower legs are in the medial wall of the precentral gyrus, with the thighs, trunk, and shoulder at the crest of the longitudinal fissure. The hand and face are in the lateral face of the gyrus. Also, the relative space allotted for the different regions is exaggerated in muscles that have greater innervation. The greatest amount of cortical space is given to muscles that perform fine, agile movements, such as the muscles of the fingers and the lower face. The "power muscles" that perform coarser movements, such as the buttock and back muscles, occupy much less space on the motor cortex.
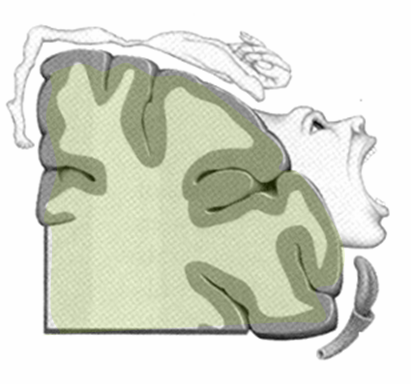
Figure 8: The above figure represents the motor homunculus man. Notice the exaggerated structures (hand and mouth). This artistic exaggeration denotes the same concept as the sensory homunculus man but with regards to motor function. The larger or more exaggerated the structure the more motor control over that structure.
The motor output from the cortex descends into the brain stem and to the spinal cord to control the musculature through motor neurons. Neurons located in the primary motor cortex are large multipolar neurons that extend from the primary motor cortex to the appropriate level in the spinal cord. This one neuron does not leave the CNS, although it does descend the spinal cord, which is why it is termed an upper motor neuron. The two descending pathways travelled by the upper motor neurons are the corticospinal tract and the corticobulbar tract. Both tracts are named for their origin in the cortex and their targets—either the spinal cord or the brain stem (the term "bulbar" refers to the brain stem as the bulb). These two descending pathways are responsible for the conscious or voluntary movements of skeletal muscles. The axons of the corticobulbar tract are ipsilateral, meaning they project from the cortex to the motor nucleus on the same side of the nervous system. Conversely, the axons of the corticospinal tract are largely contralateral, meaning that they cross the midline of the brain stem or spinal cord and synapse on the opposite side of the body. Therefore, the right motor cortex of the cerebrum controls muscles on the left side of the body, and vice versa.
The corticospinal tract descends from the cortex through the deep white matter of the cerebrum. With continued descent, upon entering the medulla, the tracts decussate. At the appropriate level of the spinal cord this upper motor neuron will synapse with lower motor neurons in the ventral grey horn of the spinal cord. The somatic nervous system provides output strictly to skeletal muscles. The lower motor neurons, which are responsible for the contraction of these muscles, are found in the ventral horn of the spinal cord. These large, multipolar neurons have a corona of dendrites surrounding the cell body and an axon that extends out of the ventral horn. This axon travels through the ventral nerve root to join the emerging spinal nerve. The lower motor neuron is also called the terminal neuron because the axon exits the spinal cord and travels to the effector thus stimulating the effector to act and cause a response. The axon is relatively long because it needs to reach muscles in the periphery of the body. The diameters of cell bodies may be on the order of hundreds of micrometers to support the long axon; some axons are a meter in length, such as the lumbar motor neurons that innervate muscles in the first digits of the feet. The axons will also branch to innervate multiple muscle fibers. Together, the motor neuron and all the muscle fibers that it controls make up a motor unit. Motor units vary in size. Some may contain up to 1000 muscle fibers, such as in the quadriceps, or they may only have 10 fibers, such as in an extraocular muscle. The number of muscle fibers that are part of a motor unit corresponds to the precision of control of that muscle. Also, muscles that have finer motor control have more motor units connecting to them, and this requires a larger topographical field in the primary motor cortex. Please see the figure below for a visual representation of the corticospinal tract.
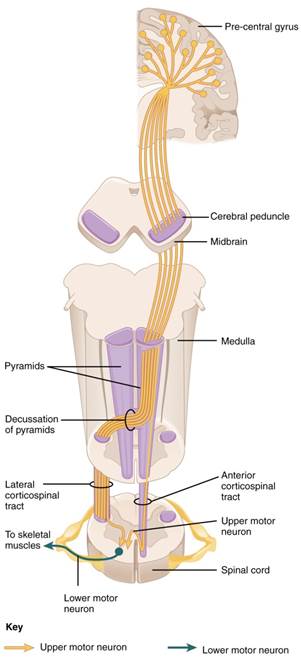
Figure 9: The major descending tract that controls skeletal muscle movements is the corticospinal tract. It is composed of two neurons, the upper motor neuron and the lower motor neuron. The upper motor neuron has its cell body in the primary motor cortex of the frontal lobe and synapses on the lower motor neuron, which is in the ventral horn of the spinal cord and projects to the skeletal muscle in the periphery.
The conscious movement of our muscles is more complicated than simply sending a single command from the precentral gyrus down to the proper motor neurons. During the movement of any body part, our muscles relay information back to the brain, and the brain is constantly sending "revised" instructions back to the muscles. The cerebellum is important in contributing to the motor system because it compares cerebral motor commands with proprioceptive feedback. The corticospinal fibers that project to the ventral horn of the spinal cord have branches that also synapse in the pons, which project to the cerebellum. Also, the proprioceptive sensations of the dorsal column system have a collateral projection to the medulla that projects to the cerebellum. These two streams of information are compared in the cerebellar cortex. Conflicts between the motor commands sent by the cerebrum and body position information provided by the proprioceptors cause the cerebellum to stimulate the midbrain, which sends corrective commands to the spinal cord.
A good example of how the cerebellum corrects cerebral motor commands can be illustrated by walking in water. An original motor command from the cerebrum to walk will result in a highly coordinated set of learned movements. However, in water, the body cannot actually perform a typical walking movement as instructed. The cerebellum can alter the motor command, stimulating the leg muscles to take larger steps to overcome the water resistance.

This chapter began by introducing reflexes as an example of the basic elements of the somatic nervous system. Simple somatic reflexes do not include the higher centers discussed for conscious or voluntary aspects of movement. Reflexes can be spinal or cranial, depending on the nerves and central components that are involved. The example described at the beginning of the chapter involved heat and pain sensations from a hot stove causing withdrawal of the arm through a connection in the spinal cord that leads to contraction of the biceps brachii. The description of this withdrawal reflex was simplified, for the sake of the introduction, to emphasize the parts of the somatic nervous system. But to consider reflexes fully, more attention needs to be given to this example.
As you withdraw your hand from the stove, you do not want to slow that reflex down. As the biceps brachii contracts, the antagonistic triceps brachii needs to relax. Because the neuromuscular junction is strictly excitatory, the biceps will contract when the motor nerve is active. Skeletal muscles do not actively relax. Instead the motor neuron needs to "quiet down," or be inhibited. In the hot-stove withdrawal reflex, this occurs through an interneuron in the spinal cord. The interneuron's cell body is located in the dorsal horn of the spinal cord. The interneuron receives a synapse from the axon of the sensory neuron that detects that the hand is being burned. In response to this stimulation from the sensory neuron, the interneuron then inhibits the motor neuron that controls the triceps brachii. This is done by releasing a neurotransmitter or other signal that hyperpolarizes the motor neuron connected to the triceps brachii, making it less likely to initiate an action potential. With this motor neuron being inhibited, the triceps brachii relaxes. Without the antagonistic contraction, withdrawal from the hot stove is faster and keeps further tissue damage from occurring.
Another example of a withdrawal reflex occurs when you step on a painful stimulus, like a tack or a sharp rock. The nociceptors that are activated by the painful stimulus activate the motor neurons responsible for contraction of the tibialis anterior muscle. This causes dorsiflexion of the foot. An inhibitory interneuron, activated by a collateral branch of the nociceptor fiber, will inhibit the motor neurons of the gastrocnemius and soleus muscles to cancel plantar flexion. An important difference in this reflex is that plantar flexion is most likely in progress as the foot is pressing down onto the tack. Contraction of the tibialis anterior is not the most important aspect of the reflex, as continuation of plantar flexion will result in further damage from stepping onto the tack. Due to the flexion of one muscle group occuring simultaneously to the extension of another muscle group, this reflex is often called the crossed flexor-extensor reflex.
Another type of reflex is a stretch reflex. In this reflex, when a skeletal muscle is stretched, a muscle spindle receptor is activated. The axon from this receptor structure will cause direct contraction of the muscle. A collateral branch of the muscle spindle fiber will also inhibit the motor neuron of the antagonist muscles. The reflex helps to maintain muscles at a constant length. A common example of this reflex is the knee jerk that is elicited by a rubber hammer struck against the patellar ligament in a physical exam. See the figure below for a representation of the reflex.
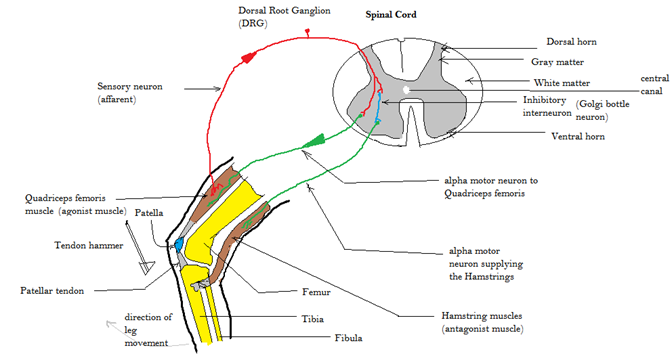
Figure 10: This figure illustrates the patellar reflex. Notice that the tendon hammar depresses the patellar tendon which pulls on the patella and quadriceps tendon. This action thereby stretches the quadriceps femoris muscle activating the muscle spindle receptors. The information is sent up the afferent neuron and integrated in the spinal cord. Notice the two motor outputs in green, one to the quadricep and the other to the hamstring. The outputs are antagonistic in nature and stimulate the quadricep while simulatenously inhibiting the hamstring thus allowing extension at the knee joint. CCBY: Amiya Sarkar
Watch this video to learn more about newborn reflexes and then answer the question below. Newborns have a set of reflexes that are expected to have been crucial to survival before the modern age. These reflexes disappear as the baby grows, as some of them may be unnecessary as they age. The video demonstrates a reflex called the Babinski reflex, in which the foot flexes dorsally and the toes splay out when the sole of the foot is lightly scratched. This is normal for newborns, but it is a sign of reduced myelination of the spinal tract in adults.
Video Companion Question
Now that we are familiar with the process of sensation it is important to address a very important concept, adaptation. Adaptation refers to the decrease in the activity of first order neurons in response to a constant stimulus. What does this mean? When you woke up this morning and were getting dressed you were very conscious of the clothes that you donning. Are they comfortable? Too hot? Too scratchy? Etc. Once you got dressed and headed on to the next part of you day you no longer paid attention to the clothing although the stimulus was still present. It is only when the stimulus changes (shirt gets stuck in a door) or your attention is called to the stimulus (as I just did by mentioning your clothing) that you become consciously aware of the stimulus again. This conscious fading of a stimulus even though the stimulus itself is still present is called adaptation. This process is vital to survival so that the cerebrum can concentrate on the most important task at hand and let the trivial stimuli fade to the background thus prevent perpetual distraction.
This concept of adaptation also illustrates another very important point; not every stimulus that reaches the cerebrum will elicit a response. With limited resources and time to act stimuli are ranked in order of importance and integrated to form a complete picture of the external environment. This allows the brain to respond efficiently to either only the imperative stimulus or to several at one time.
Though visceral senses are not primarily a part of conscious perception, those sensations sometimes make it to conscious awareness. If a visceral sense is strong enough, it will be perceived. The sensory homunculus—the representation of the body in the primary somatosensory cortex—only has a small region allotted for the perception of internal stimuli. If you swallow a large bolus of food, for instance, you will probably feel the lump of that food as it pushes through your esophagus, or even if your stomach is distended after a large meal. If you inhale especially cold air, you can feel it as it enters your larynx and trachea. These sensations are not the same as feeling high blood pressure or blood sugar levels.
When particularly strong visceral sensations rise to the level of conscious perception, the sensations are often felt in unexpected places. For example, strong visceral sensations of the heart will be felt as pain in the left shoulder and left arm. This irregular pattern of projection of conscious perception of visceral sensations is called referred pain. Depending on the organ system affected, the referred pain will project to different areas of the body (see the figure below). The location of referred pain is not random, but a definitive explanation of the mechanism has not been established. The most broadly accepted theory for this phenomenon is that the visceral sensory fibers enter into the same level of the spinal cord as the somatosensory fibers of the referred pain location. By this explanation, the visceral sensory fibers from the mediastinal region, where the heart is located, would enter the spinal cord at the same level as the spinal nerves from the shoulder and arm, so the brain misinterprets the sensations from the mediastinal region as being from the axillary and brachial regions. Projections from the medial and inferior divisions of the cervical ganglia do enter the spinal cord at the middle to lower cervical levels, which is where the somatosensory fibers enter.
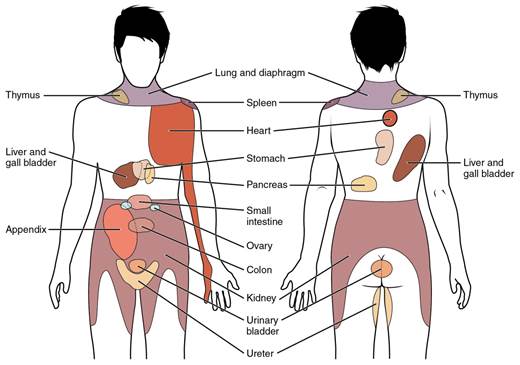
Figure 11: Conscious perception of visceral sensations map to specific regions of the body, as shown in this chart. Some sensations are felt locally, whereas others are perceived as affecting areas that are quite distant from the involved organ. CCBY: OpenStax college
Kehr's sign is the presentation of pain in the left shoulder, chest, and neck regions following rupture of the spleen. The spleen is in the upper-left abdominopelvic quadrant, but the pain is more in the shoulder and neck. How can this be?
The incorrect assumption would be that the visceral sensations are coming from the spleen directly. In fact, the visceral fibers are coming from the diaphragm. The nerve connecting to the diaphragm takes a special route. The phrenic nerve is connected to the spinal cord at cervical levels 3 to 5. The motor fibers that make up this nerve are responsible for the muscle contractions that drive ventilation. These fibers have left the spinal cord to enter the phrenic nerve, meaning that spinal cord damage below the mid-cervical level is not fatal by making ventilation impossible. Therefore, the visceral fibers from the diaphragm enter the spinal cord at the same level as the somatosensory fibers from the neck and shoulder.
The diaphragm plays a role in Kehr's sign because the spleen is just inferior to the diaphragm in the upper-left quadrant of the abdominopelvic cavity. When the spleen ruptures, blood spills into this region. The accumulating hemorrhage then puts pressure on the diaphragm. The visceral sensation is actually in the diaphragm, so the referred pain is in a region of the body that corresponds to the diaphragm, not the spleen.

Except where otherwise noted, this work by The Community College Consortium for Bioscience Credentials is licensed under a Creative Commons Attribution 4.0 International License.
Text from BioBook licensed under CC BY NC SA and Boundless Biology Open Textbook licensed under CC BY SA.
Other text from OpenStaxCollege licensed under CC BY 3.0. Modified by Alice Rudolph, M.A., Michael Ayers, M.S. and Andrea Doub, M.S. for c3bc.
Instructional Design by Courtney A. Harrington, Ph.D., Helen Dollyhite, M.A. and Caroline Smith, M.A. for c3bc.
Media by Brittany Clark, Jose DeCastro, Jordan Campbell and Antonio Davis for c3bc.
This product was funded by a grant awarded by the U.S. Department of Labor's Employment and Training Administration. The product was created by the grantee and does not necessarily reflect the official position of the U.S. Department of Labor. The Department of Labor makes no guarantees, warranties, or assurances of any kind, express or implied, with respect to such information, including any information on linked sites and including, but not limited to, accuracy of the information or its completeness, timeliness, usefulness, adequacy, continued availability, or ownership.
;