Visual Pathways
Connections between the vestibular system and the cranial nerves controlling eye movement keep the eyes centered on a visual stimulus, even though the head is moving. During head movement, the eye muscles move the eyes in the opposite direction as the head movement, keeping the visual stimulus centered in the field of view.
The connections of the optic nerve are more complicated than those of other cranial nerves. Instead of the connections being between each eye and the brain, visual information is segregated between the left and right sides of the visual field. In addition, some of the information from one side of the visual field projects to the opposite side of the brain. Within each eye, the axons projecting from the medial side of the retina decussate at the optic chiasm. For example, the axons from the medial retina of the left eye cross over to the right side of the brain at the optic chiasm. However, within each eye, the axons projecting from the lateral side of the retina do not decussate. For example, the axons from the lateral retina of the right eye project back to the right side of the brain. Therefore the left field of view of each eye is processed on the right side of the brain, whereas the right field of view of each eye is processed on the left side of the brain (Figure 7).
Segregation of Visual Field Information at the Optic Chiasm
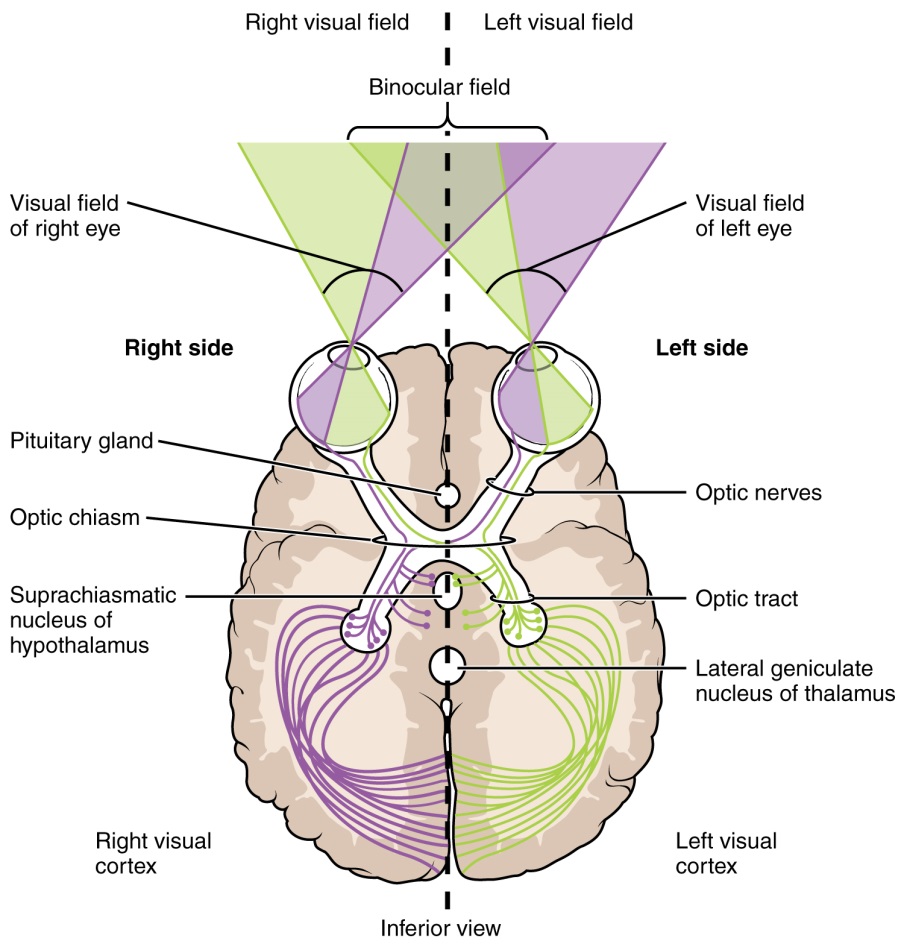
Figure 7: Contralateral visual field information from the lateral retina projects to the ipsilateral brain, whereas ipsilateral visual field information has to decussate (crossover) at the optic chiasm to reach the opposite side of the brain.
A unique clinical presentation that relates to this anatomic arrangement is the loss of lateral peripheral vision, known as bilateral hemianopia. This is different from "tunnel vision" because the superior and inferior peripheral fields are not lost. Visual field deficits can be disturbing for a patient, but in this case, the cause is not within the visual system itself. A growth of the pituitary gland presses against the optic chiasm and interferes with signal transmission. However, the axons projecting to the same side of the brain are unaffected. Therefore, the patient loses the outermost areas of their field of vision and cannot see objects to their right and left.
Extending from the optic chiasm, the axons of the visual system are referred to as the optic tract instead of the optic nerve. The optic tract has three major targets, two in the diencephalon and one in the midbrain. The connection between the eyes and diencephalon is demonstrated during development, in which the neural tissue of the retina differentiates from that of the diencephalon by the growth of the secondary vesicles. The connections of the retina into the CNS are a holdover from this developmental association. The majority of the connections of the optic tract are to the thalamus—specifically, the lateral geniculate nucleus. Axons from this nucleus then project to the visual cortex of the cerebrum, located in the occipital lobe. Another target of the optic tract is the superior colliculus.
In addition, a very small number of RGC axons project from the optic chiasm to the suprachiasmatic nucleus of the hypothalamus. These RGCs are photosensitive, in that they respond to the presence or absence of light. Unlike the photoreceptors, however, these photosensitive RGCs cannot be used to perceive images. By simply responding to the absence or presence of light, these RGCs can send information about day length. The perceived proportion of sunlight to darkness establishes the circadian rhythm of our bodies, allowing certain physiological events to occur at approximately the same time every day.
In the cerebral cortex, sensory processing begins at the primary sensory cortex, then proceeds to an association area, and finally, into a multimodal integration area. For example, the visual pathway projects from the retinae through the thalamus to the primary visual cortex in the occipital lobe. This area is primarily in the medial wall within the longitudinal fissure. Here, visual stimuli begin to be recognized as basic shapes. Edges of objects are recognized and built into more complex shapes. Also, inputs from both eyes are compared to extract depth information. Because of the overlapping field of view between the two eyes, the brain can begin to estimate the distance of stimuli based on binocular depth cues.
The centers of the two eyes are separated by a small distance, which is approximately 6 to 6.5 cm in most people. Because of this offset, visual stimuli do not fall on exactly the same spot on both retinae unless we are fixated directly on them and they fall on the fovea of each retina. All other objects in the visual field, either closer or farther away than the fixated object, will fall on different spots on the retina. When vision is fixed on an object in space, closer objects will fall on the lateral retina of each eye, and more distant objects will fall on the medial retina of either eye (Figure). This is easily observed by holding a finger up in front of your face as you look at a more distant object. You will see two images of your finger that represent the two disparate images that are falling on either retina.
These depth cues, both monocular and binocular, can be exploited to make the brain think there are three dimensions in two-dimensional information. This is the basis of 3-D movies. The projected image on the screen is two dimensional, but it has disparate information embedded in it. The 3-D glasses that are available at the theater filter the information so that only one eye sees one version of what is on the screen, and the other eye sees the other version. If you take the glasses off, the image on the screen will have varying amounts of blur because both eyes are seeing both layers of information, and the third dimension will not be evident. Some optical illusions can take advantage of depth cues as well, though those are more often using monocular cues to fool the brain into seeing different parts of the scene as being at different depths.
Retinal Disparity
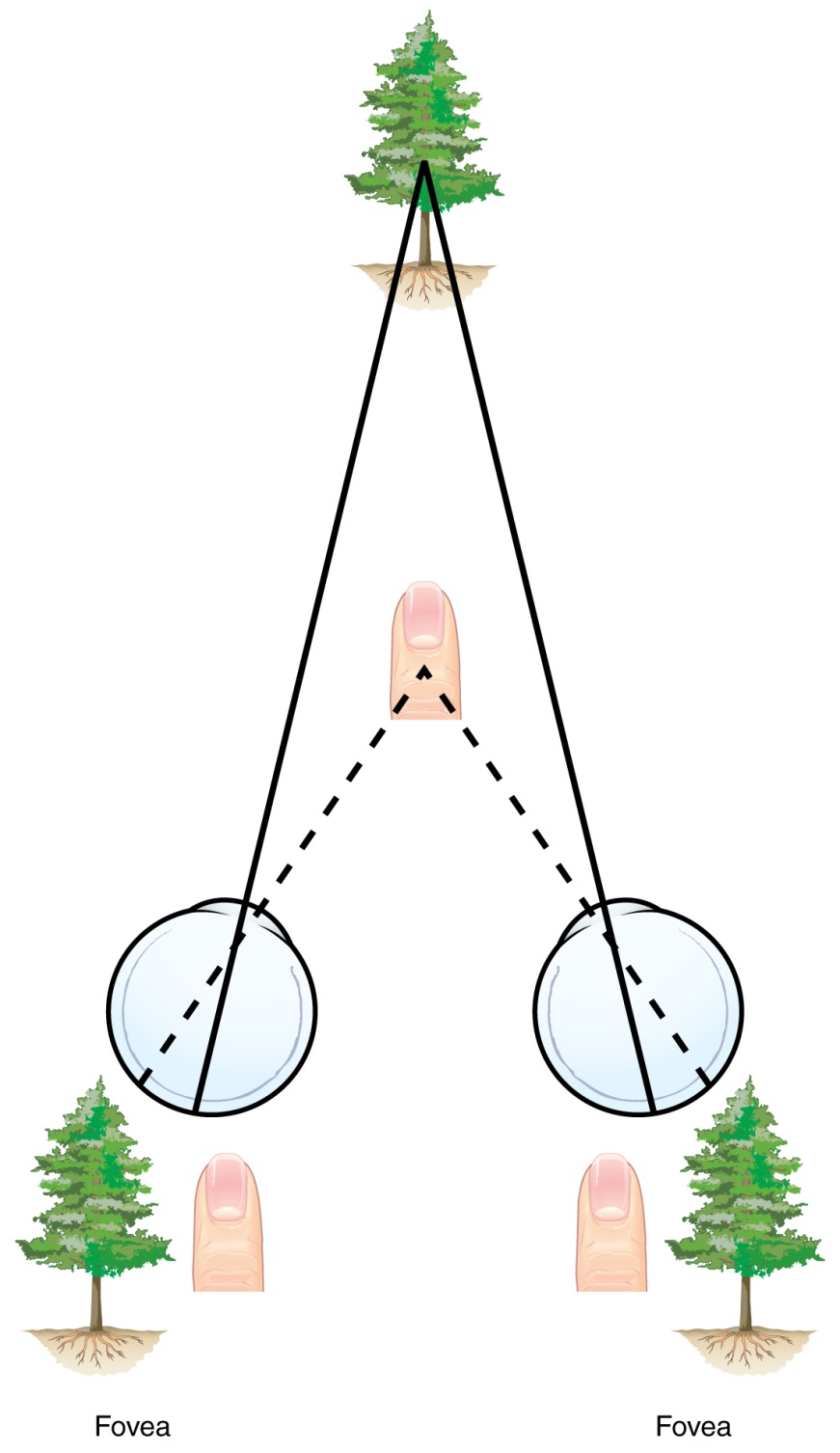
Figure 8: Because of the interocular distance, which results in objects of different distances falling on different spots of the two retinae, the brain can extract depth perception from the two-dimensional information of the visual field. If you looked at your finger in front of your eyes, with one eye closed it looks as though it is in a different place than with the other eye or with both eyes open. Using both eyes your brain interprets the appropriate depth perception.
There are two main regions that surround the primary cortex that are usually referred to as areas V2 and V3 (the primary visual cortex is area V1). These surrounding areas are the visual association cortex. The visual association regions develop more complex visual perceptions by adding color and motion information. The information processed in these areas is then sent to regions of the temporal and parietal lobes. Visual processing has two separate streams of processing: one into the temporal lobe and one into the parietal lobe. These are the ventral and dorsal streams, respectively (Figure). The ventral stream identifies visual stimuli and their significance. Because the ventral stream uses temporal lobe structures, it begins to interact with the non-visual cortex and may be important in visual stimuli becoming part of memories. The dorsal stream locates objects in space and helps in guiding movements of the body in response to visual inputs. The dorsal stream enters the parietal lobe, where it interacts with somatosensory cortical areas that are important for our perception of the body and its movements. The dorsal stream can then influence frontal lobe activity where motor functions originate.
Ventral and Dorsal Visual Streams
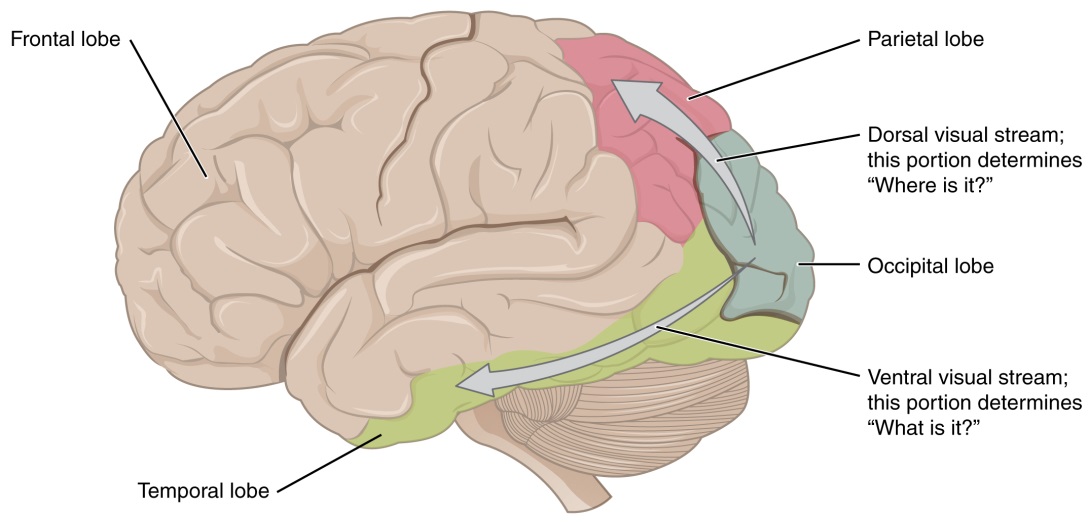
Figure 9: From the primary visual cortex in the occipital lobe, visual processing continues in two streams—one into the temporal lobe, which determines "What is it I am seeing?", and one into the parietal lobe, which determines "Where is it?"